研究报告/Research Report
准噶尔无叶豆 EsDREB2B基因在烟草中的抗逆功能分析 
2中国科学院大学, 北京, 100049
 作者
作者  通讯作者
通讯作者
豆科基因组学与遗传学, 2016 年, 第 7 卷, 第 1 篇
收稿日期: 2015年12月11日 接受日期: 2015年12月14日 发表日期: 2016年01月20日
引用格式(中文):
李士敏, 李小双, 杨红兰, 张道远, 2016, 准噶尔无叶豆EsDREB2B基因在烟草中的抗逆功能分析, 分子植物育种, 14(2): 302-308
引用格式(英文):
Li S.M., Li X.S., Yang H.L., and Zhang D.Y., 2016, Analysis on Stress-resistant Functions of EsDREB2B Gene from Eremosparton songoricum in Tobacco, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 14(2): 302-308
DREB (Dehydration Responsive Element Binding protein) 转录因子在植物响应非生物胁迫上具有重要作用,是颇具潜力的植物抗逆育种候选基因。本研究旨在通过烟草体系,验证前期从准噶尔无叶豆中克隆的EsDREB2B基因在种子萌发和成苗成长阶段的抗逆功能,并初步探究其调控的下游靶基因。结果表明:转基因株系在干旱、高盐、冷和热胁迫处理下的萌发率都高于野生型烟草,其中盐和热胁迫下萌发率差异尤为显著;两月大转基因烟草与野生型植株进行干旱、高盐、冷和热胁迫处理后,转基因较野生型植株叶片萎蔫,黄化和伤害程度较轻,在热胁迫下差异最为显著;恢复培养后,大多野生型烟草表现为枯死状态,转基因植株则叶片萎蔫减轻并开始生长,并且转基因烟草存活率显著高于野生型,均达到了野生型的3~4倍。ERD10B和Hsp70基因在四种胁迫后的转基因烟草中的表达量均较野生型高,Hsp90在热胁迫下较野生型表达量升高,而MnSOD,P5CS等基因的表达量变化不显著。以上结果表明,EsDREB2B基因能提高烟草抗干旱、高盐、冷和热胁迫的能力,是具有潜力的抗逆分子育种候选基因。
极端的环境例如干旱,盐,高温以及低温等,会严重影响植物的生长。为了适应环境,植物在生理生化及分子水平做出了响应,其中包括一些抗性基因的表达。抗性基因主要分为两种,功能基因和转录因子(Wang et al., 2003)。转录因子是一类重要的上游调控因子,通过与反式作用因子相互结合来调控下游一系列基因的表达(刘强等, 2000, 科学通报, 45(14): 1465-1474)。DREB (The dehydration-responsive element-binding proteins)作为植物特有的转录因子是目前抗逆基因研究的热点之一。
DREB转录因子属于AP2/ERF家族中的一个亚家族,由高度保守的结构域AP2和两个保守元件YRG以及RAYD组成(Agarwal et al., 2006),可以特异性的结合DRE(dehydration responsive element)顺式作用元件,序列为(TACCGACAT) (Yamaguchishinozaki and Shinozaki, 1994),诱导下游相关基因的表达,使植株从整体水平获得抵抗环境胁迫的能力。DREB转录因子可分DREB1和DREB2两类,DREB1类受低温胁迫强烈诱导,而DREB2类受干旱、高盐和热胁迫强烈诱导。目前已从大量植物中分离鉴定出DREB基因(Challam et al., 2015; Huang et al., 2008; Liu et al., 2007; Stolf-Moreira et al., 2010),并通过转化这些基因得到了抗逆性增强的转基因植株(Mallikarjuna et al., 2011; Morran et al., 2011; Zhang et al., 2009)。但是,在过表达DREB基因的同时,许多植株出现了矮化和畸形(Matsukura et al., 2010),因此寻求不仅能增加植物的抗逆性,又不会影响植株生长状态的DREB基因成为亟待解决的问题(杜洪伟等, 2008)
准噶尔无叶豆(Eremosparton songoricum (Litv.) Vass.)系多年生豆科、无叶豆属超旱生无叶灌木(张道远等, 2008),仅片断化分布于新疆古尔班通古特沙漠流动沙丘上(尹林克等, 2006)。作为西北干旱荒漠区的标志性植物种群,长期的环境压力使得该种具有抗旱、抗高温、抗风蚀沙埋等抵抗多种非生物胁迫的能力,而其中潜在的抗逆性基因鲜为报道。
前期研究中,项目组从极端抗旱植物准噶尔无叶豆中克隆得到一个新奇的天然截短型EsDREB2B基因,并在2周大的烟草幼苗中验证了该基因具有广谱的抗非生物胁迫性能(Li et al., 2014)。本研究进一步系统验证了该基因在烟草种子萌发和成苗生长两个阶段的抗逆特性,并初步探究该基因对可能下游靶基因的调控作用。研究对于进一步深入理解EsDREB2B基因的抗逆功能和抗逆机理奠定基础;同时,研究结果可直接服务于抗逆分子育种,提供遗传背景清晰、抗逆性能优良的新的基因元件。
1 结果与分析
1.1 转基因烟草的鉴定
将潮霉素筛选的T1代阳性植株移入营养土中培养一段时间,随机挑选14株待检测的转基因株系,提取叶片DNA和RNA进行PCR和RT-PCR检测。
通过PCR检测,14株转基因植物中有8株可以扩增出特异的目的条带(图1)。对这8株转基因株系进行进一步的RT-PCR检测,最终有5株扩增出特异的目的条带,分别为泳道3,4,6,8,9(图2)。挑选了泳道6,9对应的株系,分别命名为L1,L2进行进一步的抗逆性分析。
|
图1转基因烟草鉴定的PCR电泳图 |
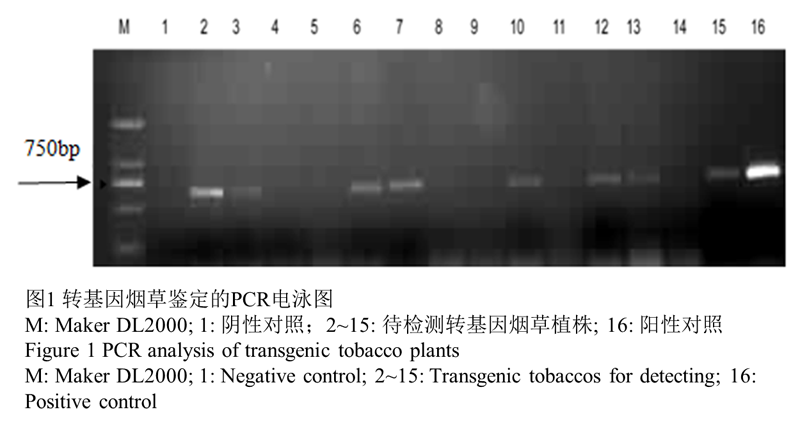
|
图2转基因烟草鉴定的RT-PCR电泳图 |
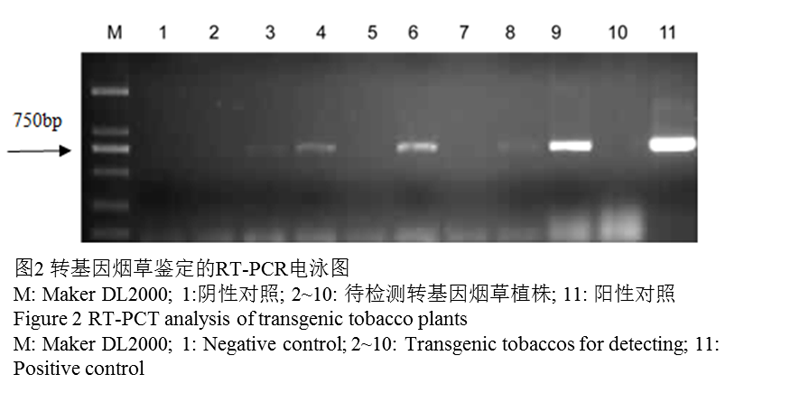
1.2 转基因烟草种子的萌发率
WT和转基因(L1和L2)T2代种子在各种胁迫下的萌发率(图3):在正常的MS培养基上,WT和转基因种子的萌发率没有明显的差异,萌发率都接近100%。在高盐和热胁迫的情况下,WT种子的萌发率分别为9%和60%,而转基因株系L1萌发率分别为29%和88%,L2为48%和98%,通过数据显示转基因株系L1和L2萌发率都显著的高于WT。在经受干旱胁迫后,转基因株系L2在萌发的数量上高于WT。除此之外,与其它DREB2类转录因子不同的是,转基因株系L1的种子在4°C条件下的萌发率为93%,表现出了明显的抗冷性,WT萌发率则为66%,显著性的低于转基因株系L1。
|
图3转基因烟草种子萌发率 |
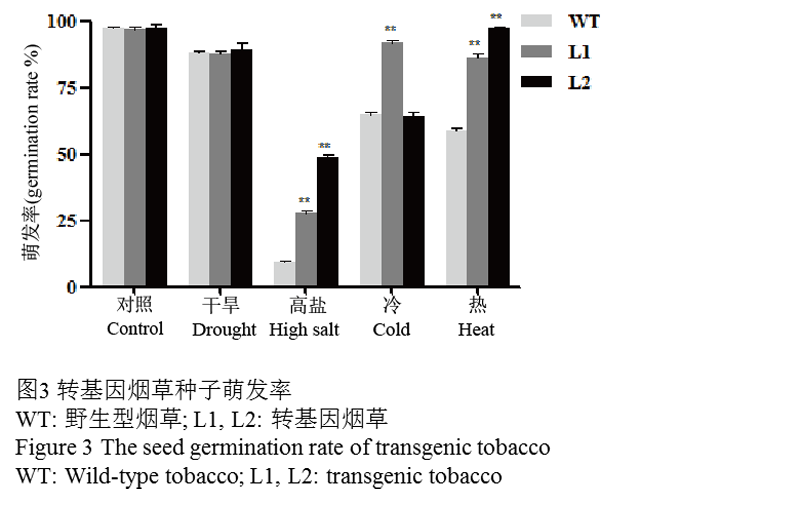
1.3 四种胁迫对WT和转基因烟草生长和存活率的影响
挑选转基因L1,L2株系和WT植株,进行干旱、高盐、冷和热胁迫,并在胁迫后复水,观察表型并统计存活率(图4)。表型观察显示(图4A):在各种胁迫处理前,转基因植株和对照植株长势相当;在干旱胁迫后,转基因和WT均呈现叶片边缘卷曲并呈暗绿色的表型,WT植株甚至出现下部叶片枯黄的症状,而转基因的植株萎蔫和受伤害程度较轻,且复水后,WT植株呈现枯死状态,而转基因植株却能成活并恢复生长;在高盐胁迫后,转基因和WT植株均呈现不同程度的萎蔫症状,然而WT整株都呈现黄化表象,转基因植株则只有下部叶片黄化,而恢复培养后,WT基本不能恢复,转基因植株虽仍有轻微萎蔫症状,但较胁迫时状态已有了一定程度的恢复;冷胁迫后,转基因植株和WT植株均呈现严重冻害的表型,植株整体严重萎蔫,叶片颜色呈墨绿色,厚而脆,恢复后,WT植株大部分死亡,而转基因植株却基本长出新叶;在热胁迫后,WT和转基因植株都出现严重的萎蔫,移至正常室温恢复生长后,WT植株基本枯死,而转基因植株则有少部分恢复正常生长。胁迫后存活率的统计结果发现(图4B):无论是转基因还是WT株系在胁迫后都出现了一定程度的死亡,然而总体上转基因植株较WT表现出了更高的存活率。在干旱和盐胁迫后,转基因植株的成活率大约是WT的3倍,转基因植株的为65%,WT为20%。而在冷和热胁迫后,转基因植株的存活率分别为60%和20%,而WT只有15%和5%,转基因植株的存活率达到了野生型的4倍。
以上实验证明,转EsDREB2B基因植株较WT具有更强的抵抗胁迫能力和更高的自我恢复能力。
|
图4两月大转基因和野生型烟草植株在干旱、高盐、冷、热胁迫下的表型和存活率 |
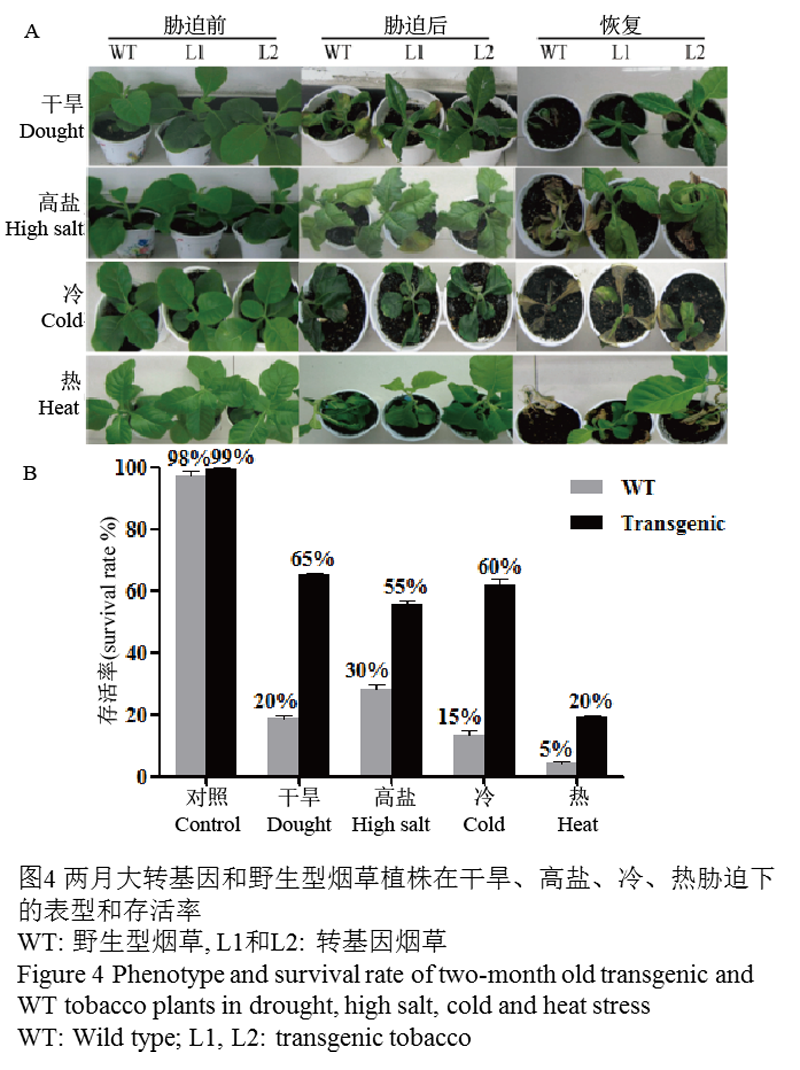
1.4 下游基因表达分析
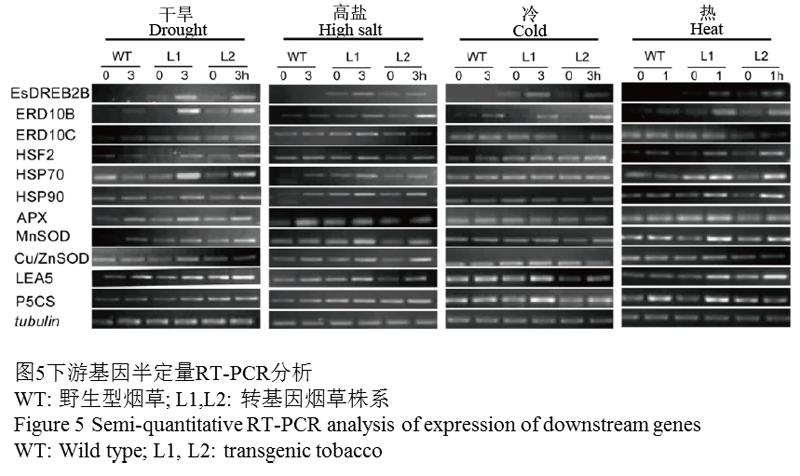
选取已经报道过的DREB基因的下游基因如脱水相关基因ERD10B、ERD10C、LEA,抗氧化相关基因APX、MnSOD、Cu/ZnSOD、CAT,脯氨酸合成关键酶基因P5CS,热激蛋白相关Hsp70、Hsp90、HSF等来初步探索EsDREB2B的可能的下游基因。结果显示(图5),在转基因烟草当中,EsDREB2B表达量在胁迫前和胁迫后均有表达;下游基因ERD10B胁迫前不表达(如冷胁迫)或者只有少量的表达(如干旱,盐和热),当胁迫3小时后,各种胁迫下的转基因植株当中ERD10B的表达量都表现为不同程度的增加;转基因植株中Hsp70除在冷胁迫后没有出现明显的表达量变化外,经过其它三种胁迫后都出现表达量上调的趋势,在干旱和热胁迫下上调最为明显;经过热胁迫后,Hsp90基因在转基因株系L1和L2中出现明显的上调。而在WT当中,无论是胁迫前还是胁迫后,基因表达量均无明显的变化趋势。以上结果说明,ERD10B,Hsp70和Hsp90不同程度的响应了EsDREB2B的表达,而其它下游基因(ERD10C、LEA、APX、MnSOD、Cu/ZnSOD、CAT、P5CS、HSF)可能不参与EsDREB2B调控的信号通路。
|
图5下游基因半定量RT-PCR分析 |
2 讨论
目前,DREB2类基因提高转基因植物抗逆性的报道较多,但都局限于某一个或两个特定胁迫抗性。通过对一些主要的DREB2基因过表达的转基因植株的抗性研究的文献整理发现:过表达AtDREB2A-CA (Sakuma, 2006),OsDREB2B;2(Matsukura et al., 2010),ZmDREB2A(Qin et al., 2007)和GmDREB2A(Mizoi et al., 2013)的转基因拟南芥显著增强了抗干旱、热胁迫的能力,但抗高盐的能力则没有提高。过表达CAP2 (Shukla et al., 2006; Shukla et al., 2009)、PgDREB2A (Agarwal et al., 2010)和LcDREB (Ban et al., 2011)基因提高了转基因植物的干旱、高盐和热胁迫的耐受能力。而过表达EsDREB2B却能全面提高转基因烟草的抗旱、高盐、冷和热的能力。
同时,有些DREB2基因的转基因植物虽然提高了逆境胁迫下的抗性,但是正常生长条件下,转基因植物表现出严重的生长抑制或缺陷(Matsukura et al., 2010; Qin et al., 2007)。虽然RD29诱导型启动子的应用,改善了对正常情况下转基因植物长势的影响(如OsDREB2B; 2),但综合抗性的提高却一直是个难题。EsDREB2B和CAP2基因是目前极少数能够同时提高转基因植物三大非生物胁迫抗性(干旱、高盐和热胁迫),且正常生长条件下不会造成转基因植物生长缺陷的DREB2类基因。此外,转EsDREB2B基因的烟草还显示出了一定的抗冷能力。而如上所提到的能广泛响应干旱、高盐、冷、热和脱落酸胁迫的OsDREB2B基因,其转基因植株仅表现出了抗干旱和热胁迫的能力。
DREB作为一个逆境胁迫相关的转录因子基因,一个重要的特征就是它能调控下游许多基因的表达,从而快速,高效的启动植物体内一系列的抗逆调控途径。EsDREB2B转基因烟草植株在胁迫后除了ERD10B,Hsp70等少数几个基因表现出在干旱胁迫后是上调表达的,其余大部分基因如APX、MnSOD、P5CS,Lea、Hsp90、HSF等在转EsDREB2B基因烟草里上调不明显,而这些基因却在AtDREB2A (Sakuma, 2006),LbDREB (Ban et al., 2011),PgDREB2A (Agarwal et al., 2010)的转基因植株里被证明强烈诱导表达。这些结果初步说明了EsDREB2B的下游调控基因可能与非截短型的DREB2基因的有所不同,更加细致和大规模的深入分析需要开展。
因此,EsDREB2B不以牺牲植物生物量为成本,是目前报道的极少数抗逆广谱的作物抗逆分子育种的候选基因。同时也为进一步的探讨沙漠植物准噶尔无叶豆的抗逆分子机理提供了分子水平上的理论依据。
3 材料与方法
3.1 试验材料
实验用烟草种子为Petite Havana SR1。pMD19-T-EsDREB2B质粒由本实验室保存。农杆菌菌株LBA4404由中科院新疆理化与技术研究所肖向文研究员提供。植物表达载体pCAMBIA1301由新疆大学生命科学与技术学院张富春教授提供。
3.2 试验方法
3.2.1 真核表达载体的构建与鉴定
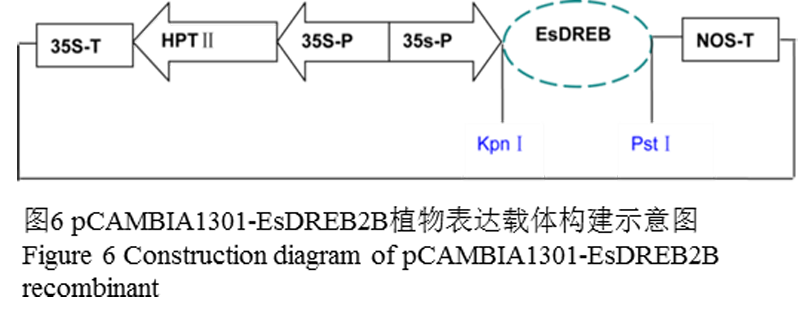
用KpnI和PstI分别酶切pMD19-T-EsDREB2B质粒载体和植物真核表达载体pCAMBIA1301,分别回收酶切产物,连接,转化大肠杆菌细胞。用EsDREB2B基因ORF引物ORF-f:5’-ATGAGTGCAACTTGCATGCA-3’和ORF-r:5’-AGACAATGAAGGATCGGAGG-3’做质粒PCR,同时以KpnI和PstI做质粒双酶切来确定重组质粒pCAMBIA1301-EsDREB2B的成功构建。pCAMBIA1301-EsDREB2B植物表达载体构建的示意图(图6):
|
图6 pCAMBIA1301-EsDREB2B植物表达载体构建示意图
|
3.2.2 农杆菌介导的烟草遗传转化
烟草的转化采用叶圆盘法(Fisher and Guiltinan, 1995)。烟草种子用浓度75%酒精浸泡30 s,1%NaClO浸泡10 min,然后用无菌水冲洗5遍,接种于MS培养基上,16h光照,28°C培养30d。将剪好的叶盘(直径约为5cm)叶片放入OD600值为0.6~0.8农杆菌侵染液,10 min后28°C黑暗条件下进行共培养。4~6d后,放于选择分化培养基上培养,每10~15d继代一次。选择培养2~3周后,待叶盘周围分化出抗性芽,且抗性芽长至2~3cm时,切下插入到选择生根培养基中,进行生根培养。待有5~7条幼根且长3~4cm的时候,移出炼苗4~6d,再转入沙土培养。
3.2.3 转基因烟草的鉴定
将获得的转基因烟草的种子进行潮霉素抗性筛选,并对具有抗性的植株进行DNA水平的PCR检测和RNA水平的RT-PCR检测。植株总DNA的提取采用CTAB法(Porebski et al., 1997),总RNA用TAKARA公司RNAiso试剂提取。以野生型(WT)烟草总DNA或cDNA为阴性对照,以阳性重组质粒pCAMBIA1301-EsDREB2B为阳性对照。挑选DNA和RNA水平均能检测到特异目的条带的株系为转基因阳性植株。每代种子进行如上筛选,直至筛选获得T2代种子。
3.2.4 转基因烟草的萌发率分析
转基因两个株系(L1和L2)的T2代种子和野生型烟草种子各150粒平铺在各种MS胁迫培养基中进行萌发实验。用MS+200mM/L甘露醇并置于正常生长温度28°C培养20d来模拟干旱胁迫;用MS+150mM/LNaCl并置于正常生长温度28°C培养20d来模拟盐胁迫(于瑞等,2013);将MS培养基放入4°C培养4d用来模拟冷胁迫;置于37°C培养5d模拟热胁迫处理。各种胁迫结束后,统计转基因烟草和野生型烟草的萌发率。
3.2.5 转基因烟草的抗逆性分析
将在MS培养基上正常生长至两周大的转基因株系T3代幼苗LI,L2以及WT植株移栽到营养土的花盆中,正常生长至两个月大时,开始实施各种胁迫。转基因各个株系和野生型烟草各20株用于此实验。对于干旱胁迫,用10% PEG6000浇两月大苗,每株(每盆)每次浇灌200mL,每3d浇一次,持续12d后正常浇水进行复水培养14d;对于盐胁迫,用300mM/L NaCl溶液浇灌幼苗,每株每次200mL,每3d浇一次,持续12d后进行复水14d;对于冷胁迫,将两月大苗置于温度4°C培养箱中进行冷锻炼,正常浇水(每3d浇灌一次,每株每次200 mL自来水),培养6d后移至温度-20°C胁迫1h,之后转移至正常温室(28°C)中进行恢复培养14d。对于热胁迫,将两月大苗置于温度45°C培养箱中胁迫12h后移至正常温室(28°C)中进行恢复培养14d。
胁迫前,胁迫后和恢复三个阶段均拍照观察植株表型,并统计在各个胁迫情况下转基因株系和野生型植株的存活率。
3.2.6下游基因表达量分析
对胁迫后的两月大转基因和WT植株进行RNA提取,然后挑选与植物脱水、抗氧化和热激胁迫等相关的10个被报道过的DREB下游基因,用烟草TUB2作为内参基因,进行半定量RT-PCR实验,来探讨EsDREB2B基因可能的下游调控基因。以Genbank下载的各个基因序列为模板,用Primer premier5设计各基因引物,实验中所用各参考基因的登录号,功能及引物序列等信息见(表1)。
|
表1 Primer information for semi-quantitative RT-PCR |
.png)
作者贡献
李士敏执行本研究的试验设计、试验研究与论文的撰写;李小双完成后期试验数据的整理与图片的处理;杨红兰参与试验的指导;张道远负责试验设计的指导以及论文审校与修改等。
致谢
本研究由新疆维吾尔自治区重大专项项目(201330122-3)资助。
Agarwal P., Agarwal P.K., Joshi A.J., Sopory S.K., and Reddy M.K., 2010, Overexpression of PgDREB2A transcription factor enhances abiotic stress tolerance and activates downstream stress-responsive genes, Molecular biology reports, 37(2): 1125-1135
Agarwal P.K., Agarwal P., Reddy M.K., and Sopory S.K, 2006, Role of DREB transcription factors in abiotic and biotic stress tolerance in plants, Plant cell reports, 25(12): 1263-1274
Ban Q., Liu G., and Wang Y., 2011, A DREB gene from Limonium bicolor mediates molecular and physiological responses to copper stress in transgenic tobacco, Journal of plant physiology, 168(5): 449-458
Challam C., Ghosh T., Rai M., and Tyagi W., 2015, Allele mining across DREB1A and DREB1B in diverse rice genotypes suggest a highly conserved pathway inducible by low temperature, Journal of Genetics, 94(2): 231-238
Du H.W., Chen F., and Xiao G.Y., 2008, Study on DREB Transcriptional factors for improvement of corp drought tolerance, Shengwu Jishu Tongbao(Biotechnology bulletin), 6: 1-6(杜洪伟, 陈芬, 肖国樱, 2008, 提高作物耐旱性的DREB转录因子研究进展, 生物技术通报, 6: 1-6)
Fisher D.K., and Guiltinan M.J., 1995, Rapid, Efficient Production of Homozygous Transgenic Tobacco Plants with Agrobacterium tumefaciens: A Seed-to-Seed Protocol, Plant Molecular Biology Reporter, 13(3): 278-289
Huang B., Jin L., and Liu J.Y., 2008, Identification and characterization of the novel gene GhDBP2 encoding a DRE-binding protein from cotton (Gossypium hirsutum), Journal of plant physiology, 165(2): 214-223
Li X.S., Zhang D.Y., Li H.Y., Wang Y.C., Zhang Y.M., and Wood A.J., 2014, EsDREB2B, a novel truncated DREB2-type transcription factor in the desert legume Eremosparton songoricum, enhances tolerance to multiple abiotic stresses in yeast and transgenic tobacco, BMC Plant Biology, 14: 44
Liu N., Zhong N.Q., Wang G.L., Li L.J., Liu X.L., He Y.K., and Xia G.X., 2007, Cloning and functional characterization of PpDBF1 gene encoding a DRE-binding transcription factor from Physcomitrella patens, Planta, 226(4): 827-838
Mallikarjuna G., Mallikarjuna K., Reddy M.K., and Kaul T., 2011, Expression of OsDREB2A transcription factor confers enhanced dehydration and salt stress tolerance in rice (Oryza sativa L.), Biotechnology letters, 33(8): 1689-1697
Matsukura S., Mizoi J., Yoshida T., Todaka D., Ito Y., Maruyama K., Shinozaki K., and Yamaguchi-Shinozaki, K., 2010, Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Molecular genetics and genomics : MGG, 283(2): 185-196
Mizoi J., Ohori T., Moriwaki T., Kidokoro S., Todaka D., Maruyama K., Kusakabe K., Osakabe Y., Shinozaki K., and Yamaguchi-Shinozaki K., 2013, GmDREB2A;2, a canonical DEHYDRATION-RESPONSIVE ELEMENT-BINDING PROTEIN2-type transcription factor in soybean, is posttranslationally regulated and mediates dehydration-responsive element-dependent gene expression, Plant physiology, 161(1): 346-361
Morran S., Eini O., Pyvovarenko T., Parent B., Singh R., Ismagul A., Eliby S., Shirley N., Langridge P., and Lopato S., 2011, Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors, Plant biotechnology journal, 9(2): 230-249
Porebski S., Bailey L.G., and Baum B.R., 1997, Modification of a CTAB DNA Extraction Protocol for Plants Containing High Polysaccharide and Polyphenol Components, Plant Molecular Biology Reporter, 15(1): 8-15
Qin F., Kakimoto M., Sakuma Y., Maruyama K., Osakabe Y., Tran L.S., Shinozaki K., and Yamaguchi-Shinozaki K., 2007, Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L., The Plant journal, 50(1): 54-69
Sakuma Y., Maruyama K., Osakabe Y., Qin F., Seki M., Shinozaki K., and Yamaguchi-Shinozaki K., 2006, Functional Analysis of an Arabidopsis Transcription Factor, DREB2A, Involved in Drought-Responsive Gene Expression, The Plant Cell, 18(5): 1292-1309
Shukla R.K., Raha S., Tripathi V., and Chattopadhyay D., 2006, Expression of CAP2, an APETALA2-family transcription factor from chickpea, enhances growth and tolerance to dehydration and salt stress in transgenic tobacco, Plant physiology, 142(1): 113-123
Shukla R.K., Tripathi V., Jain D., Yadav R.K., and Chattopadhyay D., 2009, CAP2 enhances germination of transgenic tobacco seeds at high temperature and promotes heat stress tolerance in yeast, The FEBS journal, 276(18): 5252-5262
Stolf-Moreira R., Medri M.E., Neumaier N., Lemos N.G., Brogin R.L., Marcelino F.C., de Oliveira M.C., Farias J.R., Abdelnoor R.V., and Nepomuceno A.L., 2010, Cloning and quantitative expression analysis of drought-induced genes in soybean. Genetics and molecular research: GMR, 9(2): 858-867
Wang W., Vinocur B., and Altman A., 2003, Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance, Planta, 218(1): 1-14
Yamaguchishinozaki K., and Shinozaki K., 1994, A Novel Cis-Acting Element in an Arabidopsis Gene Is Involved in Responsiveness to Drought, Low-Temperature, or High-Salt Stress, Plant Cell, 6(2): 251-264
Yin L.K., Tan L.X., and Wang B., eds., 2006, Rare Endangered Endemic Higher Plants in Xinjiang of China, Xinjiang Scientific and Technical Publishing House, Urumq, pp.74-75(尹林克, 谭丽霞, 王兵, 2006, 新疆珍稀濒危特有高等植物, 新疆:新疆科学技术出版社, pp.74-75)
Yu R., Hu Y., Huang X.X., Shen Z.D., Han Y.S., Deng S.R., Zhao R.,Shen X., and Chen S.L., 2013, Overexpression of Kandelia candel C2H2 Zinc-Finger Protein Gene KcZFP Enhanced Salinity Tolerance in Tobacco Plant, Genomics and Applied Biology, 32(2): 149-158(于瑞, 胡月, 黄旭新, 申泽丹, 韩彦莎, 邓澍荣, 赵瑞, 沈昕和陈少良, 2013, 过表达秋茄C2H2型锌指蛋白基因KcZFP提高烟草耐盐性, 基因组学与应用生物学, 32(2): 149-158)
Zhang D.Y., Ma W.B., Shi X., Wang J.C., Wang X.Y., 2008, Distribution and Bioecological Characteristics of Eremosparton songoricum a Rare Plant in Gurbantunggut Desert, Zhongguo Shamo(Journal of desert research), 28: 430-436(张道远, 马文宝, 施翔, 王建成, 王习勇, 2008, 准噶尔无叶豆的地理分布、群落学特征及生物生态学特性, 中国沙漠, 28: 430-436)
Zhang Y., Chen C., Jin X.F., Xiong A.S., Peng R.H., Hong Y.H., Yao Q.H., and Chen J.M., 2009, Expression of a rice DREB1 gene, OsDREB1D, enhances cold and high-salt tolerance in transgenic Arabidopsis, BMB reports, 42(8): 486-492